
| Home Page | Overview | Site Map | Index | Appendix | Illustration | About | Contact | Update | FAQ |
|
All animals digest their food, carry on gas exchange, excrete waste, circulate nutrient and waste products to and from the cells, coordinate their movements, protect themselves, and reproduce and disperse the species. The more complex animals have organ systems to carry out these functions; in simple animals, these functions sometimes are carried out by specialized tissues. All these functions can be found in the Human Organ Systems as shown in Figure 01a or in Table 10-01, Topic 10. Although endocrine system does not exist in invertebrates, neurosecretory systems are widespread. Such systems probably exist in all phyla. Work to date has demonstrated the important role of neurosecretion in growth and reproduction of many model systems. |
Figure 01a Human Organs [view large image] |
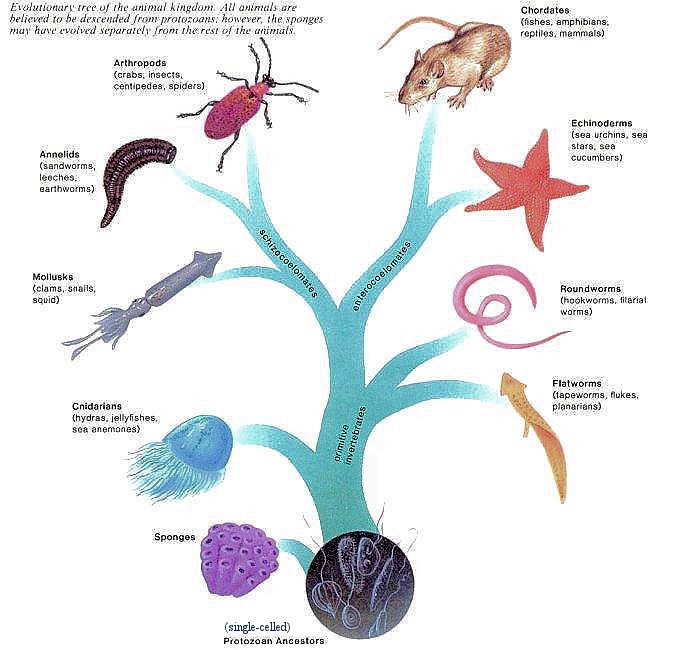 |
All phyla of animals had evolved by the beginning of the Paleozoic Era some 540 million years ago. The evolutionary tree of animals in Figure 01b indicates that animals are the descendants of protozoans - perhaps in a colonial form whose cells differentiated into various types of cells. The evolutionary tree of animals resembles a tree with 2 main branches. The animal phyla located on the main trunk of the tree are referred to as the primitive invertebrates, and the animals of the main 2 branches include the advanced invertebrates and the vertebrates. Invertebrates lack a dorsal backbone, while vertebrates have a backbone made up of vertebras. A study of the evolution of animals reveals that the most complex animals have the most advanced features as listed in Table 01 below. Classification of animals therefore is based on type of body plan, symmetry, number of germ layers, level of organization, type of body cavity, and presence or absence of segmentation. |
Figure 01b Evolutionary Tree of Animals |
| Features | Most Primitive | Primitive | Advanced | Most Advanced |
|---|---|---|---|---|
| Body Plan | None | Sac plan | Tube-within-tube plan | + specialization of parts |
| Symmetry | None | Radial | Bilateral | + cephalization§ |
| Germ layers | None | 2 | 3 | 3 |
| Level of organization | None | Tissue | Organ | Organ system |
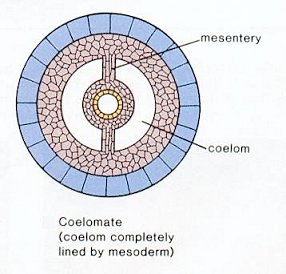
| Body cavity | Diploblastic | Acoelomate | Pseudocoelom | True coelom |
| Segmentation | Nonsegmented | Nonsegmented | Segmented | + specialization of parts |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}