 |
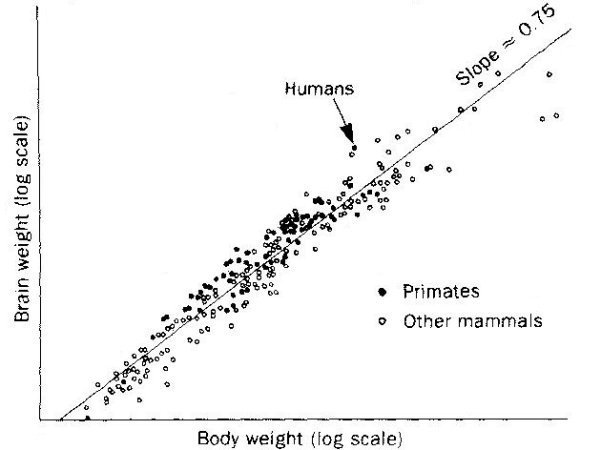 |
There is a more general 1/4 power-law applicable to many physicological variables y as shown in Figures 04, 05, and Table 01. The general form of the power-law is y  Mb, where b is a multiple of 1/4. It seems that all these physicological variables have something to do with the nutrient distribution networks and the dimensional dependence. For example, the life span is proportional to the linear dimension L, and the heart beat rate is related to the inverse L-1. The other variables simply follow the same 3/4 power-law or L3 as for the case of metabolic rate. Mb, where b is a multiple of 1/4. It seems that all these physicological variables have something to do with the nutrient distribution networks and the dimensional dependence. For example, the life span is proportional to the linear dimension L, and the heart beat rate is related to the inverse L-1. The other variables simply follow the same 3/4 power-law or L3 as for the case of metabolic rate.
|





 5x10-3 watts/gm. Whereas the R
5x10-3 watts/gm. Whereas the R 