
| Home Page | Overview | Site Map | Index | Appendix | Illustration | About | Contact | Update | FAQ |
|
Organisms have evolved to co-ordinate their activities according to the Earth's rotation, its revolution around the Sun, and the Moon's revolution around the Earth. Table 10-02 is a summary of the various types of biological rhythm. Biological cycles were thought to be passive, driven by environmental cues such as changes in light and temperature. It is only in the late 1990s that an internal-clock more or less independent of external environment was identified at molecular level. Figure 10-16a depicts the daily rhythm of a typical individual. It shows that there is a cyclic variation for most of the biological functions, which attain their high or low point at certain time each day. |
Figure 10-16a Circadian Cycle [view large image] |
| Celestial Movement | Length of Cycle | Name | Chronobiological Name | Example |
|---|---|---|---|---|
| Earth's Rotation | 12.4 hours | tidal | CIRCA TIDAL | mollusc feeding |
| Earth's Rotation | 24 hours | daily | CIRCADIAN | sleep-wake cycle |
| Moon's Revolution | 29 days | monthly | CIRCA LUNAR | menstrual cycle |
| Earth's Revolution | 365 days | yearly | CIRCANNUAL | rutting in deer |
 |
called the suprachiasmatic nuclei (SCN), which are linked to the pineal gland. Figure 10-16b shows the mechanism of the circadian clock in the brain. The ganglion cells in the retina of the eye operate independently of the rods and cones, which mediate vision. They track fluctuations in light but are far less responsive to sudden changes or low intensity. That sluggishness befits a circadian system. It would be no good if watching fireworks or going to a movie tripped the mechanism. These cells send information about brightness and duration to the SCN, which then dispatches the information to the parts of the brain and body that control circadian processes. In response to daylight, the SCN emits signals (red arrow) that stop another brain region - the paraventricular nucleus - from producing a message that would ultimately result in melatonin's release by the pineal gland. After dark, however, the SCN releases the brake, allowing the paraventricular nucleus to relay a "secrete melatonin" signal (green arrows) |
Figure 10-16b Circadian Clock [view large image] |
through neurons in the upper spine and the neck to the pineal gland. |
 |
 |
their expression peaked in the heart at different hours than in the liver. Circadian rhythms take days and sometimes weeks to adjust to a sudden shift in day length or time zone. A new schedule of light will slowly reset the SCN clock. But the other clocks may not follow its lead. The body is not only lagging; it is lagging at a dozen different places and hence the phenomenon of "jet lag", which doesn't last, presum-ably because all of those different drummers eventually sync up again. Seasonal rhythms in many animals such as hibernation, migration, molting and mating may |
Figure 10-17a Clock Mechanism [view large image] |
Figure 10-17b Biological Clock [view large image] |
also be regulated by the circadian clock, which is equipped to keep track of the length of days and nights. |
 |
To be of any biological use, the above-mentioned 24-hour molecular cycle has to be turned into a signal that can regulate physiology and behavior. For example, the SCN shows a 24-hour rhythm in electrical activity that drives output rhythms such as melatonin from the pineal. It is found that within the SCN there is a group of what are called "clock-controlled genes" or ccgs, which are driven by the molecular oscillation. This rhythmic transcription seems to involve the same basic elements that drive the molecular feedback loop, with a CLOCK/BMAL1 complex binding to the E-box element instead of CLOCK/CYCLE. One of these ccgs is the gene for the neuropeptide arginine vasopressin (avp). In the SCN, avp has a strong circadian rhythm in both its mRNA and protein abundance. It increases the electrical activity of many SCN neurons. SCN also releases avp in a rhythmic manner to alter the activity of cells outside the SCN. Further researches indicate that the molecular basis of the clock in plants, fungi, and bacteria is different from that in mammals and insects. It seems to suggest that the biological clock may have evolved multiple times during the course of evolution. In summary, although different sets of genes seem to generate the clock in animals, plants, fungi and bacteria, they use the same fundamental mechanism - an "auto-regulative negative feedback loop" involving several genes. These genes give rise to a message (mRNA) and a protein that may cycle in a circadian manner. It acts either directly or indirectly as a transcription factor, inhibiting its own gene expression. |
Figure 10-17c Clock in Organs [view large image] |
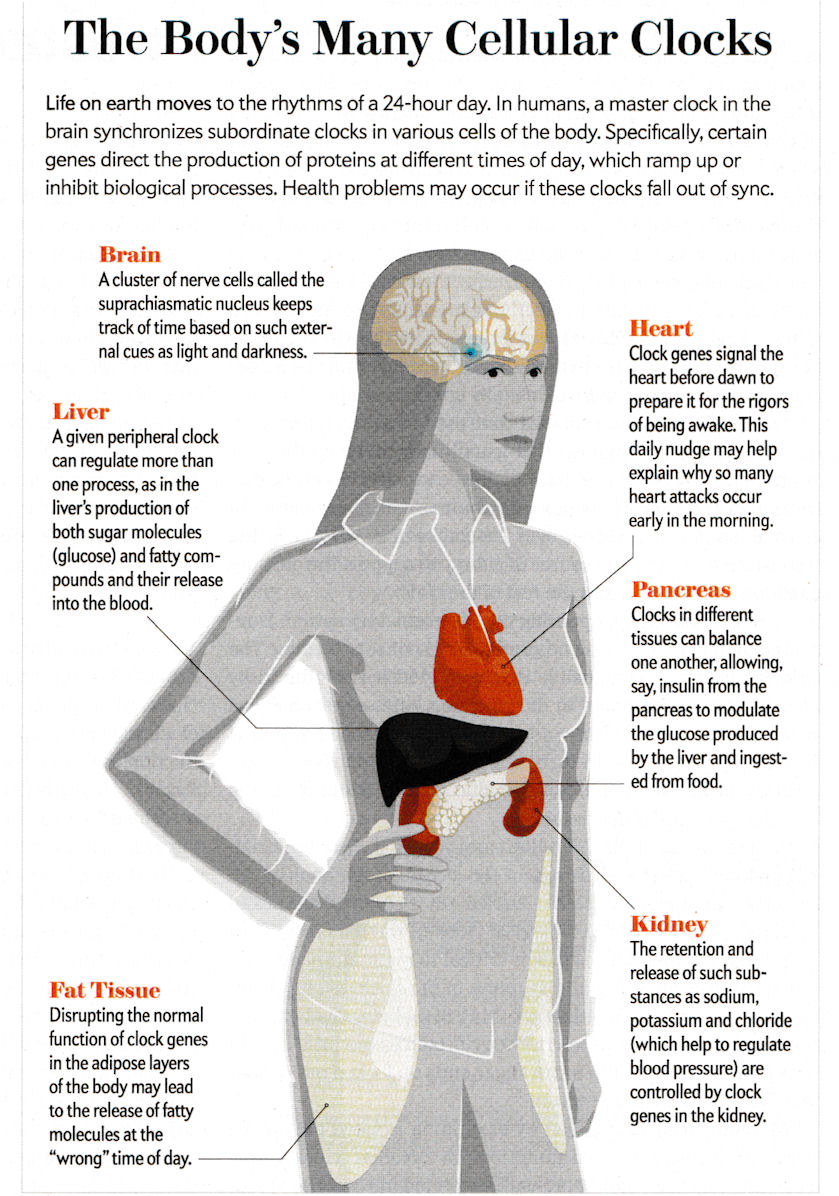
It is found recently (in the 2010s) that there are regional clock genes within many organs that control its activity (Figure 10-17c). Disruption the working of these genes can cause various health problems as obesity, diatbetes, depression etc. |
 |
 |
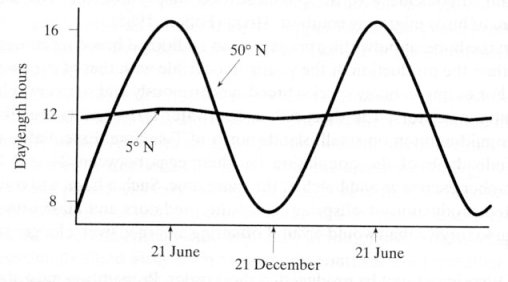
slight hint of seasonal change by the commercial contrivances of Christmas sales and chocolate Easter eggs. However, animals in the wild still depend on the ability to cope with the seasonal change for successful reproduction of progeny by carefully regulating their annual breeding. They have to survive the harsh winter every year with migration or hibernation (Figure 10-17e). Some native birds and mammals in high latitude adopt the strategy |
Figure 10-17d Daylength |
Figure 10-17e Migration and Hibernation [view large image] |
of free-running rhythms of sleep and wake allowing them to forage whenever physical conditions are favourable. |
 |
In birds, daylength is detected by deep-brain photoreceptors (DBP) and measured by a circadian clock in the SCN. Daylength information then regulates the activity of GnRH neurons in the hypothalamus. The rest of the process is similar to that for the mammals. The pineal and eye, so important in the circadian system of many birds, are not required for the photoperiodic regulation of reproduction. |
Figure 10-17f Regulation of Reproduction [view large image] |
{kind=link}