
| Home Page | Overview | Site Map | Index | Appendix | Illustration | About | Contact | Update | FAQ |
|
While engineering relies on a blueprint, architect, engineer and construction workers - all of them outside the finished product and involving human intelligence; bio-processing includes self-assembly of components (such as protein, RNA, DNA) via interaction between residual charges and adoptive self-organization in response to external circumstances. It's a continuous process that would have problem with disruption, rearrangement or piecemeal implementation. The process occurs naturally without any cue from a human elaboration. Figure 01 illustrates the different processes pictorially. |
Figure 01 Engineering vs Bio-processing |
 |
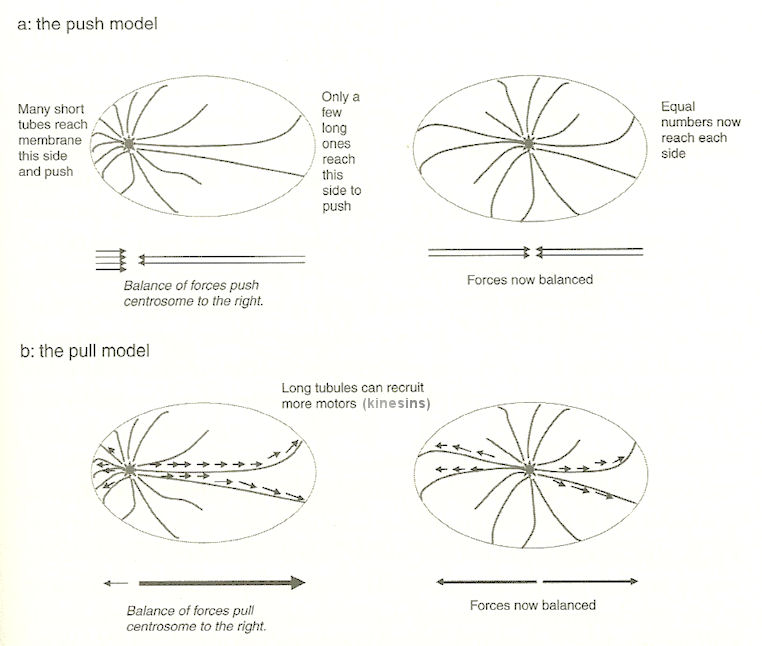 |
Both the centrosome and chromosome are divided into mother-daughter pair. The microtubules bind to a special site in the chromosomes and pull them apart (Figure 02). The binding of microtubules to the chromsomes and fresh molecules uses a trial and error approach in finding the right match. Therefore, it is expensive in terms of energy, but it is automatic. It seems to be very complicated, actually it is the feed back behavior of each individual component that gives rise to the complex from the simple. This process of first cell division is common to all cell replications by the name of mitosis. Meiosis involves further processing to produce gametes. See "Inner World of the Cell" for more information about "microtubule" and "molecular motors". |
Figure 02 Microtubule [view large image] |
Figure 03 Microtubules in Action [view large image] |
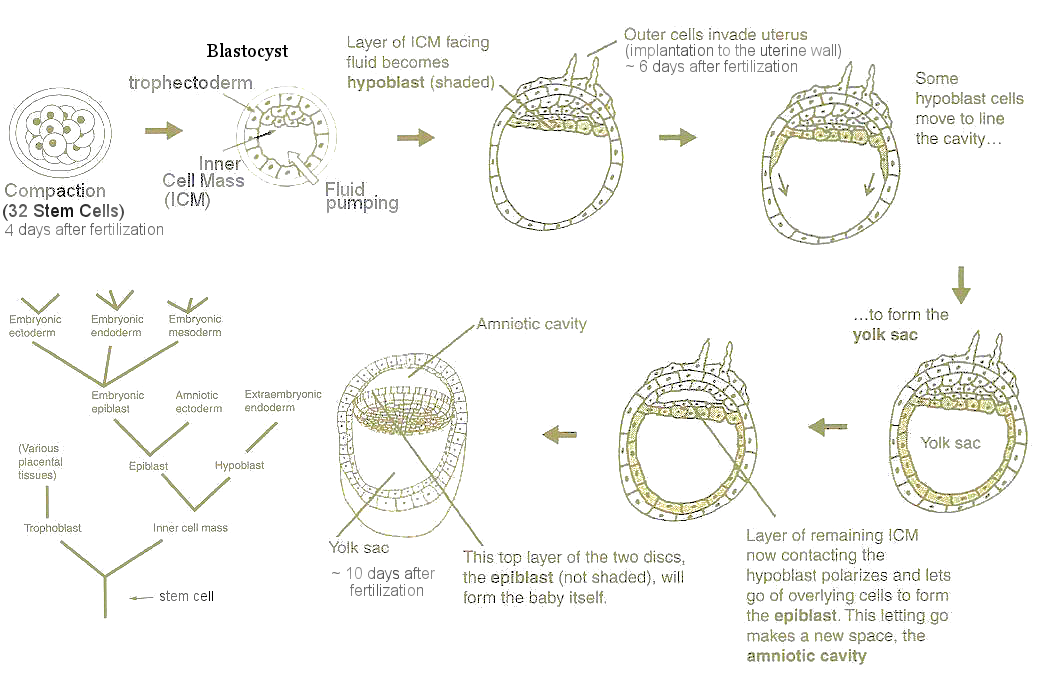 |
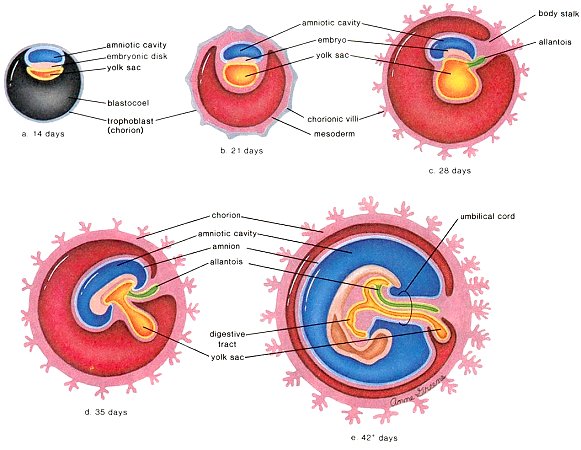
inner cell mass (ICM) moves to another end (Figure 04). The layer of ICM facing the fluid now receives another geometric cue to specialize into hypoblast cells, which spread around to form the yolk sac (providing nourishment and also functioning as internal circulation). The remaining ICM collaspes toward the hypoblast layer to form the epiblast, which is destinated to be the base of next life (Figure 04 which also shows some of the specialized cells from the stem cell). The process also creates the amniotic cavity, where fetal development occurs (see chicken egg in which the yolk sac and amnotic cavity are ready-made, and mammalian fetus which is the main subject of interest here). |
Figure 04 Spatial Cue |
 |
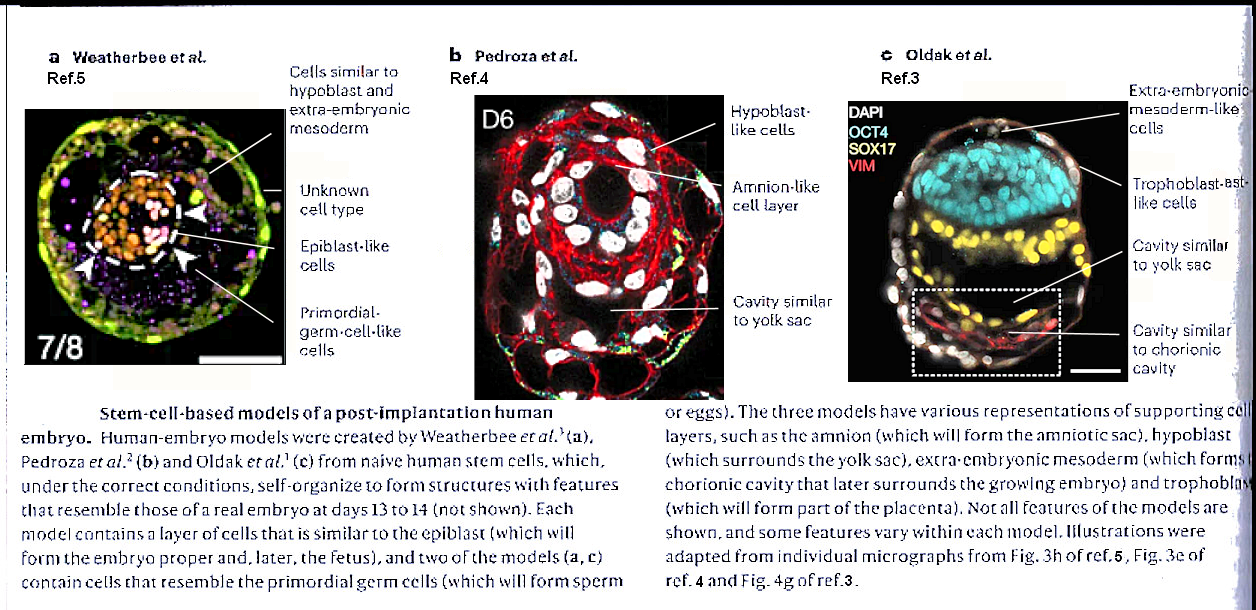
In order to bypass technical, ethical � and often legal limits in the studying of early embryo, labs around the world have made blastocyst models as show in Figure 04a in various forms according to requirement (see "Human embryo models are getting more realistic � raising ethical questions", which also shows the early development process). | |
Figure 04a Lab Made's |
 |
 |
 |
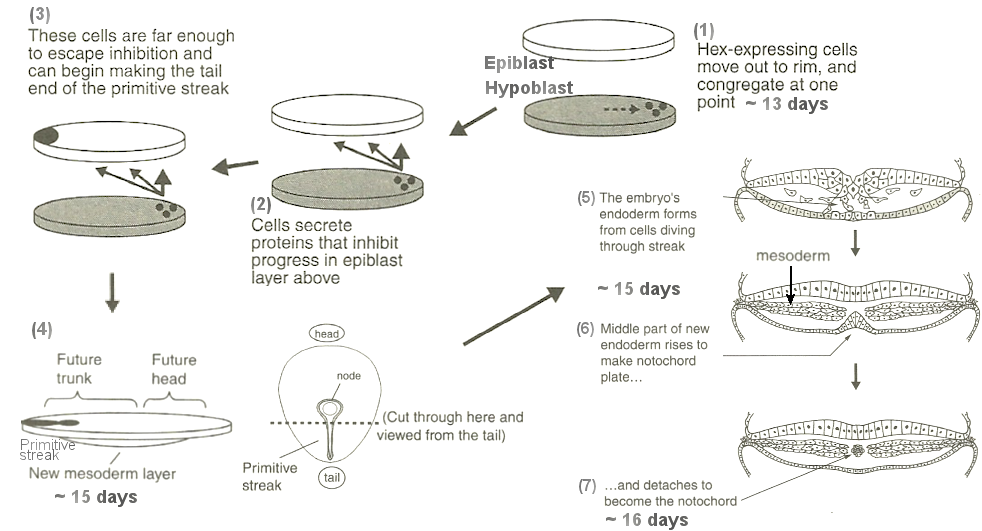
system is evolved further and further away from equilibrium in a state which requires energy infusion to sustain. It is not clear what triggers the expression of the genes that leads to all these changes. But the processes are well documented. Figures 05a and 05b depict the steps toward the laying down of the notochord from the two layers of epiblast and hypoblast (blast = form, epi = over, hypo = under). Figure 06 shows the body plan based on the three germ layers. |
Figure 05a Schematic Body Plan Development [view large image] |
Figure 05b Body Plan |
Figure 06 Germ Layers |
Followings are some elaborations on the 7 steps. |
 |
* Safety Profile: Because the body has not undergone significant changes associated with pregnancy, the risk of complications from such early interventions is very low. Medical methods used at this stage, such as emergency contraceptives, have been extensively studied and are considered safe with a well-known safety profile. | |
Figure 05 Initial 7 Days |
In summary, interventions before implantation are generally considered safe because they do not disrupt an established pregnancy and are typically associated with minimal risk. |
 |
 |
 |
 |
|
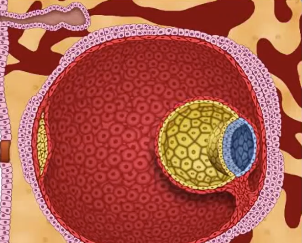
Figure 05d Embryo Day 4-6 |
Figure 05e Embryo Day 6-13 |
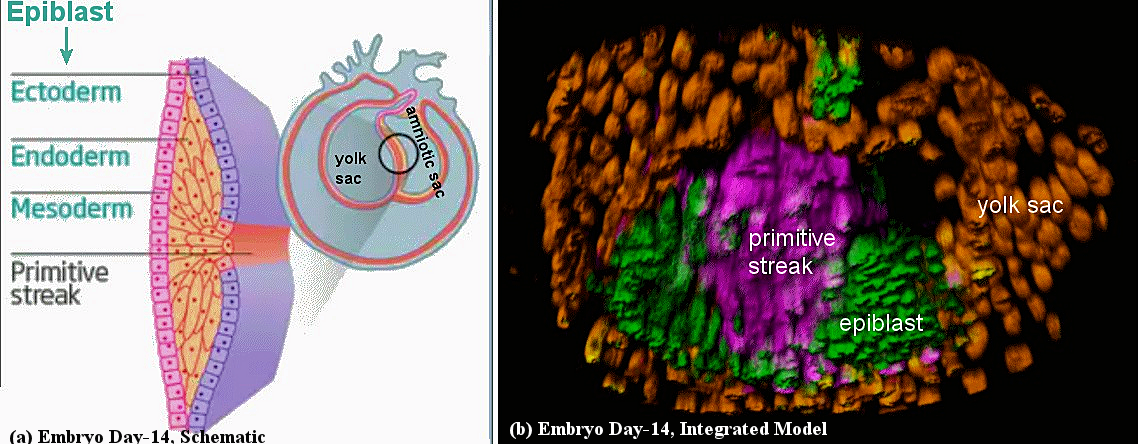
Figure 05f Embryo Day 14 [view large image] |
Figure 05g |
 |
16�19 days post-fertilization and that was undergoing a developmental process called gastrulation. During this process, a layer of cells called the epiblast gives rise to three 'germ' layers (the ectoderm, mesoderm and endoderm), establishing the body plan. At this stage, the embryo contains two cavities: a cavity formed by the amniotic ectoderm overlying the embryonic disc, and the yolk sac (surrounded by the extra-embryonic mesoderm) on the other side of the disc. The gastrulating embryo is dissected (by consent of its owner undergoing termination of her pregnancy) into three parts, a statistical analysis is used to �group� the dissociated cells according to the similarity of their gene-expression profiles. Using this approach, the cell type that make up the various embryonic structures are identified, as well as the primitive haematopoietic cells that give rise to a rudimentary blood system, and Primordial Germ Cells (PGCs - future sperm cells; not shown in Figure 05c); but shown below by a mammalian equivalence : |
Figure 05c Embryo at 16-19 Days |
 |
 |
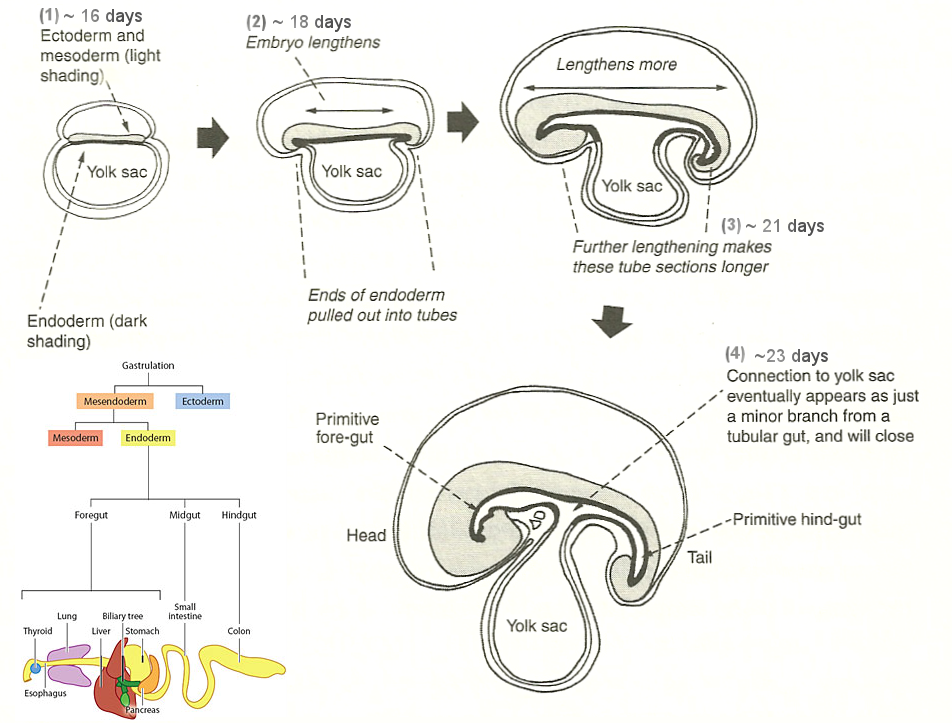 |
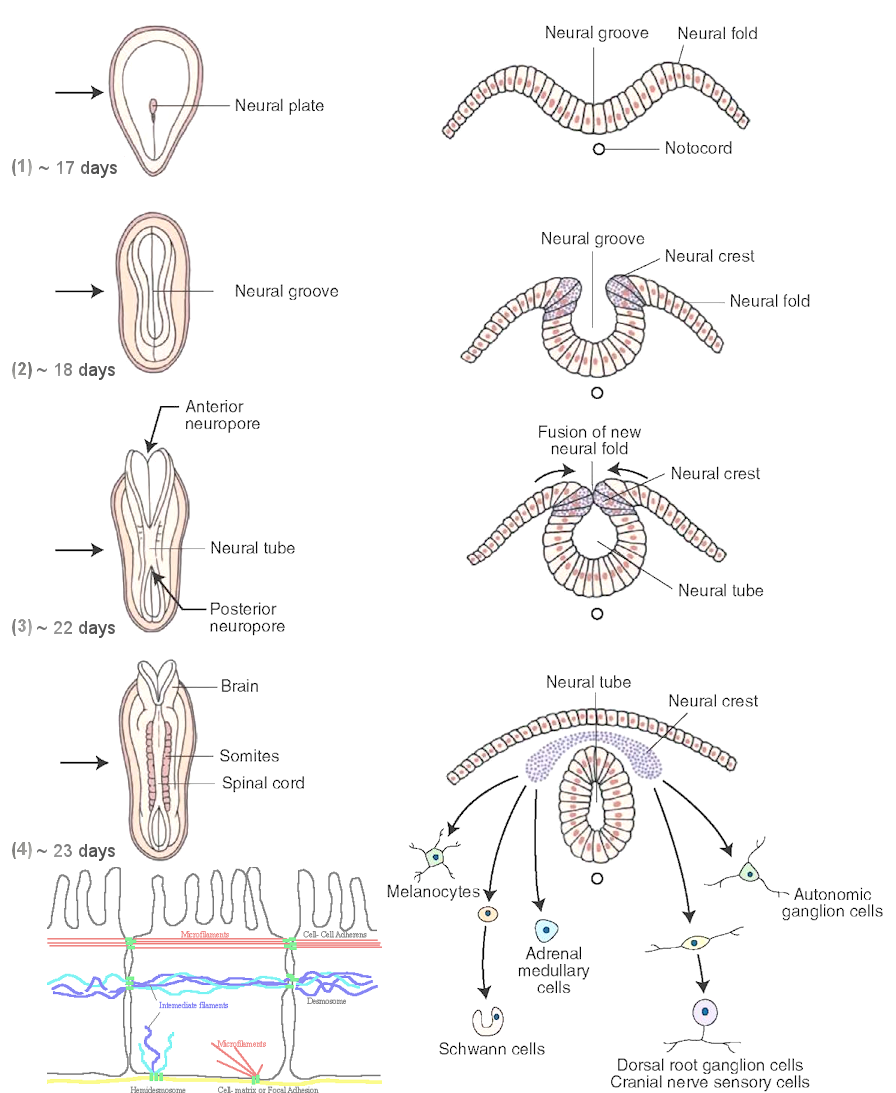
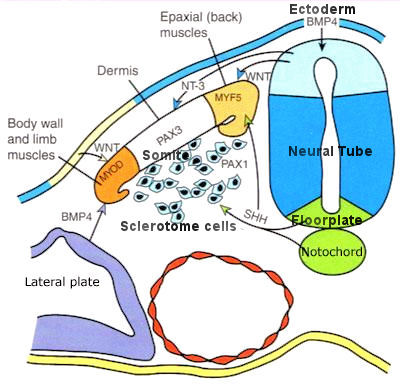
The vertebrates have two important tubing inside the body. The neural tube that contains the nerves running from the brain to the tail end, and the gut tube that retrieves nutrient from food and repels the waste at the end. Their formation is achieved either from the local change of individual cell that propagates via a continuous mechanical network or just by the growth of the body. It is the microfilaments that helps to buckle the central axis of the ectoderm to form the neural tube. While the gut tube is formed by the overgrown of the endoderm. These processes are shown in Figures 07 and 08. A brief description is provided in the followings. |
Figure 07 Neural Tube [view large image] |
Figure 08 Gut Tube |
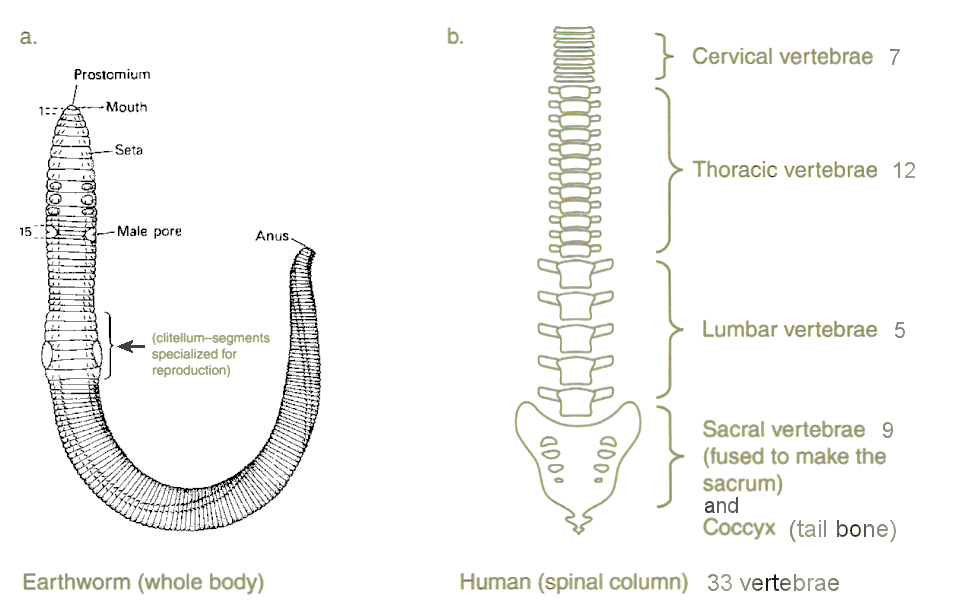 |
animals such as the earthworm (Figure 09a, a ventral view). Each segment is self-sufficient in physiological terms, having its own primitive lungs, kidneys, nervous system, etc. Human seems to be less obvious, the segmentation is concealed inside in the form of spinal column (Figure 09b). Starting from the top two vertebrae, which enables their possessor to nod and shake the head, there are 5 more in the neck, 12 in the chest with ribcage (to protect the lungs, heart, and other internal organs of the thorax; in some animals, especially snakes, ribs may provide support and protection for the entire body), 5 lumbar vertebrae to support the weight, and 9 sacrum and tail bone with a ring to protect the pelvic region. |
Figure 09 Segmentation |
 |
vertebrae, muscles of the trunk, and a few other body parts. The formation of these somites does not happen all at once, they are developed in a sequence from the neck of the embryo towards the tail (see Figure 10b, 10a is a live example of a zebrafish embryo) with some spacing in between; otherwise, its owner would have to move rigidly - cannot turn, bend, and perform any of those athletic movements. Since the embryo has now grown to a scale for which a single signal concentration gradients becomes more error prone, a different mechanism is used to generate the smites. The un-committed mesoderm cells keep sending out signal (in the form of FGF protein) to inhibit the making of new somite while the last somite produces another signaling molecule, retinoic acid, to counteract the FGF effect. |
Figure 10 Somite Development [view large image] |
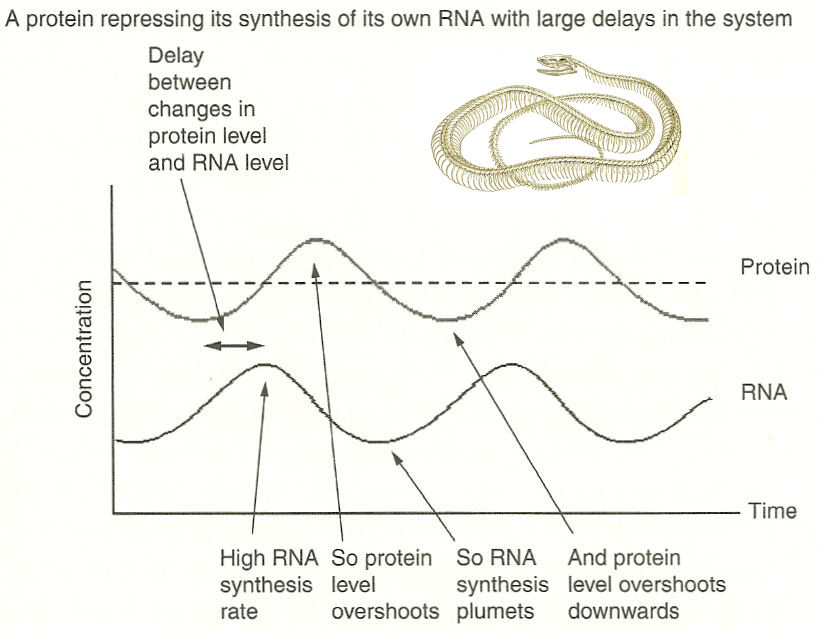 |
model it is assumed that a protein would send off signal to repress its synthesis by inhibiting the very RNA that produces the protein with some time delay. As shown in Figure 11, this pattern of under- and over-shooting repeats in cycles, making a simple oscillator or clock. The segment can be large or small depending on the rate of the clock (Figure 10c). For example, the clock in snake embryo runs a lot faster than those in mice or human, resulting in a large number of rib bones (see inert in Figure 11). In humans, a new somite forms every 6 hours or so. The actual detail is not yet fully clear, it seems to be the combination of the FGF proteins, the retinoic acids, the somite clock and tailward growth which enables the embryo to organize itself in a regular, large-scale pattern based on only local rules. |
Figure 11 Somite Clock |
 |
 |
There is very strong evidence, from other animals, that our remote ancestors had just one HOX cluster as shown in Figure 12a for the urchins, flies, ... etc. During the long history of evolution, it had been copied twice making a total of four clusters in tetrapods by the names of HOXA, HOXB, HOXC, HOXD. There are 13 variants of HOX genes all in a fixed sequence, however, each cluster would not have the whole complement. Some HOX genes may have been deleted and others mutated within each cluster over the eon. Researches in the late 1990's indicated that some lineages such as the teleost (ray-finned) fish made even more copies up to eight (Figure 12a shows six of those found). |
Figure 12a Hox Genes |
Figure 12b Hox Gene Activation |
 |
The cellular clock explanation becomes more complicated when a study compares how rapidly proteins were created and destroyed in the skin cells of 12 mammals, ranging from golden hamsters that barely live 4 years, to humans, to bowhead whales that can live 200 years. It finds a strong inverse correlation with lifespan: longer-lived species had slower turnover of proteins, i.e., the clock rate for protein turnover is slower for larger animals (Figure 13,d). Researchers are now searching for an unifying cause for these findings (see original article "These cellular clocks help explain why elephants are bigger than mice, Nature News, 27 April 2021"). |
Figure 13 Cellular Clocks |
Instead of an unifying view, the clock rate in the embryonic development could be dictated by the Hes7 or Hox genes (ultimately); while the clock rate for protein turnover is related to the "Kleiber's Law", which links the size of the animal to the circulation of nutrients and wastes, i.e., the metabolic rate R  M3/4, where M is the mass of the animal. M3/4, where M is the mass of the animal.
|
 |
A very important function of cells is the ability to secrete proteins to deliver chemical messages and to perform a task upon receiving such message, e.g., generating another chemical message or changing to another cell type. Since by the laws of physics, message from such small source as a typical cell (with a dimension about 0.001 cm) cannot go very far (with range ~ 0.005 cm), it is essentially a local action between adjacent blocks of cells. But the same kind of mechanism can be used repeatedly all over, the concentration gradient can also be used to induce different actions according to the amount. In organogenesis, different chemical message and different concentration would produce different type of cell or tissue as demonstrated in the following example in patterning the neural tube and somite to neuron, bone, muscle, and skin. |
Figure 14 Organogenesis |
 |
Cell Movement - Normally, cells are tightly packed with adhesive protein E-cadherin. Cell migration starts by lowering the E-cadherin level in the EMT (epithelial-mesenchymal transition) process by which epithelial cells lose their cell polarity and cell-cell adhesion, and gain migratory and invasive properties to become mesenchymal stem cells; these are multipotent stromal cells that can differentiate into a variety of cell types. The reverse process, MET (Mesenchymal-epithelial transition) are critical for development of many tissues and organs in the developing embryo, and numerous embryonic events such as those in gastrulation (Figuge 15, left panel). EMT has also been shown to occur in wound healing, in organ fibrosis and in the initiation of metastasis for cancer progression. |
Figure 15 Cell Movement [view large image] |
 |
like the GPS (Global Positioning System), which guides the vehicle through a network of streets to arrive at its destination. The neural crest cells migration to form various cell types is a good example to illustrate the process. Figure 16 right panel shows a few migration routes, more specific cell types or organs are displayed in the left panel, also see Figure 07. |
Figure 16 Cell Migration |
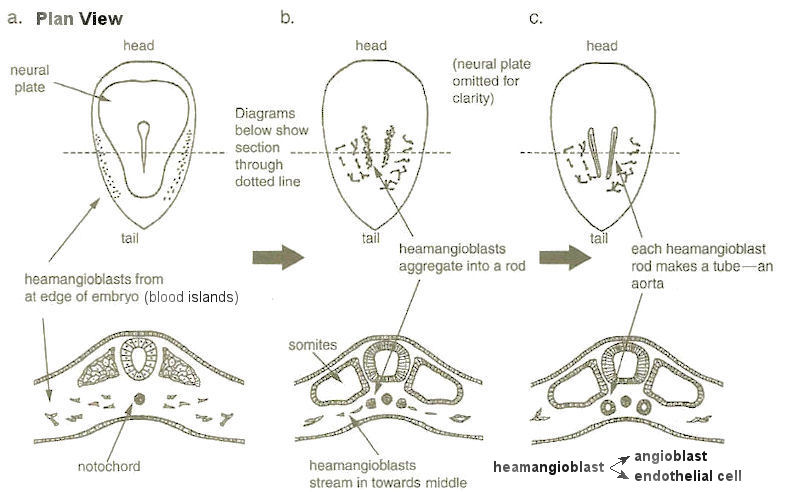 |
(Figure 17b). The cells aggregate into two solid rods on each side. The rods become hollowed out later by cell-suicide. The SHH proteins secreted by the notochord prevent them from merging into one (Figure 17c). The process is applicable to the formation of both the arteries and veins. The only difference is that the veins has larger bore and thinner wall; the concentration gradient of the SHH proteins is responsible for the two types. Blood cells are made from some of the cells in the wall of the aorta on receiving signals from gut tube and neural tube. The signallings are combined in such a way that not all the cells in the wall are converted into blood cells. The actual blood cells are made in the newly developed liver. |
Figure 17 Vasculogenesis |
 |
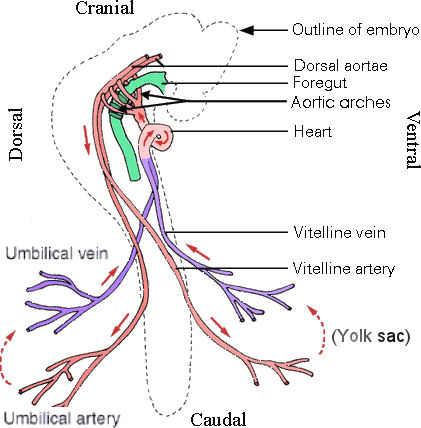 |
|
Figure 18 Heart Tube |
Figure 19 Circulation, Embryonic |
Figure 18 shows the end result of the heart tube formation process, and Figure 19 depicts the complete embryonic circulation system. |
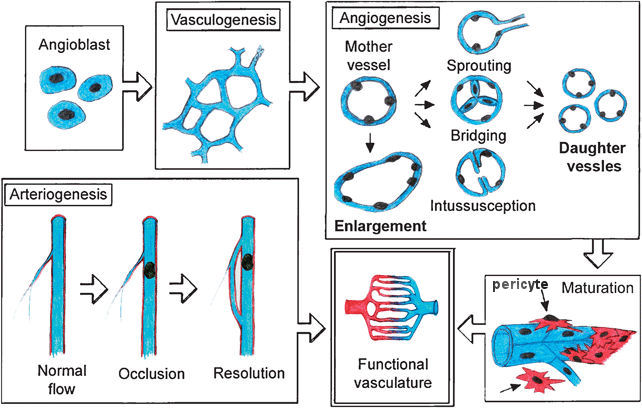 |
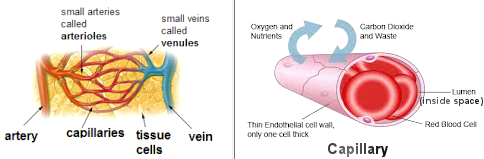
of the vessel. The generation of capillaries is vital to the health of the embryo as well as to all adults. Whenever a cell is in need of oxygen or nutrient, it sends out a signal for help. Capillaries will grow to such location to deliver the supply. The capillary wall is a one-layer endothelium that allows lipophilic (oil loving), hydrophilic (water loving) molecules and gas to pass through without the need for special transport mechanisms. It allows bidirectional diffusion depending on osmotic gradients (diffusion of fluids through membranes or porous partitions). The capillaries can also repair themselves by the process of angiogenesis. The distressed cell eventually dies off if no capillaries come to its rescue. |
Figure 20 Angiogenesis |
 |
 |
tube is controlled by signals coming from the surrounding, mesoderm-derived cells. For example, the lung buds start to form by pushing out from the gut tube. In fish, this gives rise to the swim bladder. In mammals it is heavily modified : it branches repeatedly to form a large tree-like structure. They eventually become the airways of the lungs. In all cases, it is the endoderm-derived cells, which make the tubes in the organ, while the mesoderm-derived cells compose into the rest of the solid tissue. Figure 21 also shows the beginning days for the development of the organs. |
Figure 21 Gut Organs |
Figure 22 Mesoderm Organs |
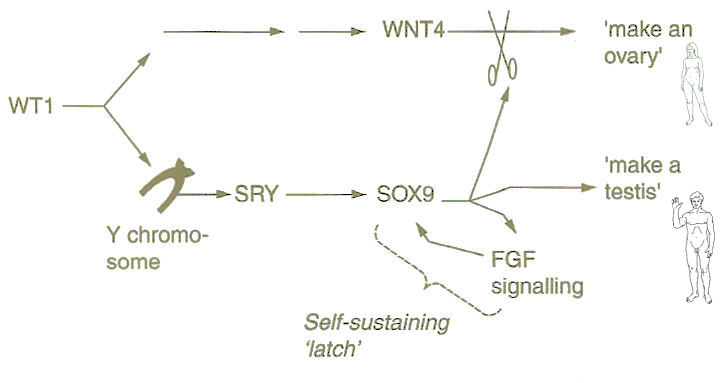 |
 |
undergo the process of meiosis to become sperms (cells in haploid state with either X or Y chromosome) or eggs (cells in haploid state with only X chromosome) depending on whether further development is following the male or female path (Figure 23). In boys, meiosis does not begin until puberty but in girls all of the germ line cells begin meiosis almost as soon as the overy begins developing. Every egg that a woman can ever have is already in meiosis when she is born. The determination of sex is governed by the gonad, which begins to make a protein called WT1 at about day 42. |
Figure 23 Sex Determination |
Figure 24 Genitals, Difference |
See "Sex and Death" for a philosophical discourse on mortal life. |
| Substance (level) | Male | Female |
|---|---|---|
| Sex Chromosome Pair (1) | XY | XX |
| SRY Protein (1) | Yes | No |
| WNT4 Protein (1) | No | Yes |
| Mullerian Duct (2) | Eliminated | Develops to oviducts, uterus, upper vagina |
| Wolffian Duct (2) | Becomes sperm transport duct | Eliminated |
| Genital (3) | Penis and scrotum | Vagina and labia |
| Mammary Glands (3) | Vestiged | Presence |
| Bones and Muscles (3) | Larger | Smaller |
| Hair (3) | More | Less |
 |
The two pairs of limbs start out as small projections underneath the ectoderm that covers the outside of the embryo. The proliferation is driving by signals from the mesoderm of the trunk. They switch on the production of the WNT proteins, which in turn induce the FGF signalling family. The FGF is the driving force to move the limb bud called "Progress Zone" outward (Figure 25a). Further development of the limb is explained by two different models as outlined below. |
Figure 25 Development of Limbs [view large image] |
 |
 |
crawling with the growth cone (Figure 26). The direction of growth is specified by the adhesion of the substratum. The branching tends to follow a stickier path. Other guidance cues work by signalling to the molecular machinery that assembles the growth cone's leading edge. Since the movement of these migrating cells excesses the growth of the embryo, they all piled up at the wall to become the meningeal layers of the CNS (Figure 27). |
Figure 26 Growth Cone |
Figure 27 Meninges [view large image] |
 |
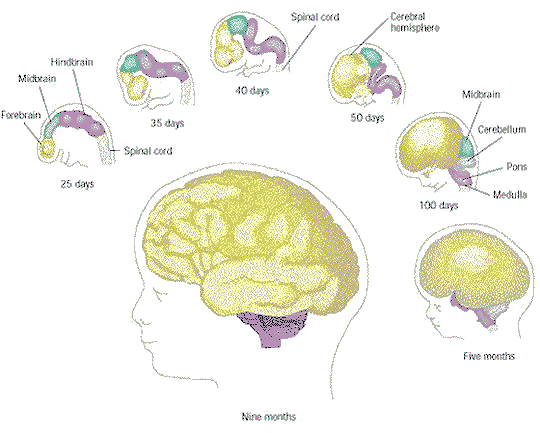 |
to cram even more surface area into the available space (Figure 29). The process follows the basic principle of chemical signalling over and over, getting more and more complicated. This is the physical base of mentality provided by nature, the owner of such system has much to learn to become a viable being after birth (see "Nervous System" for the mature version of CNS, PNS, ANS, ENS, etc.). |
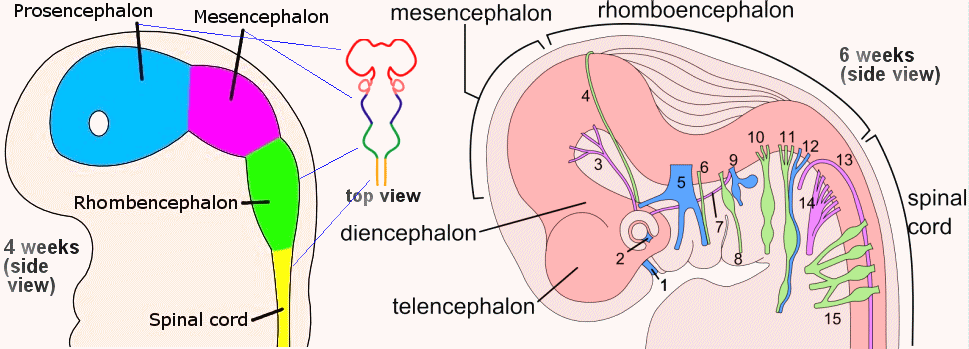
Figure 28 Embryo Brain at 4 and 6 Weeks |
Figure 29 Development, Brain [view large image] |
 |
first time at the birth canal. Delivery by caesarean section delays the contact until it is handled by the mother (see "Immune System" for the different forms of immunity). Since the adaptive immune system is acquired, the practice of good hygiene in developed countries has been accompanied by an increase in the incidence of diseases such as asthma, which reflect an immune system out of balance and inclined to be excited by harmless substances such as dust, animal hair, and pollen (Figure 30). |
Figure 30 Asthma |
BTW, It is found that infants delivered by cesarean section exhibit a distinct microbiome (the micro-ecosystems on and in our body) more colosely resembles the composition of the mother's skin. |
 |
| ||
Video 01 Early Development, |
Video 02 General Development, |
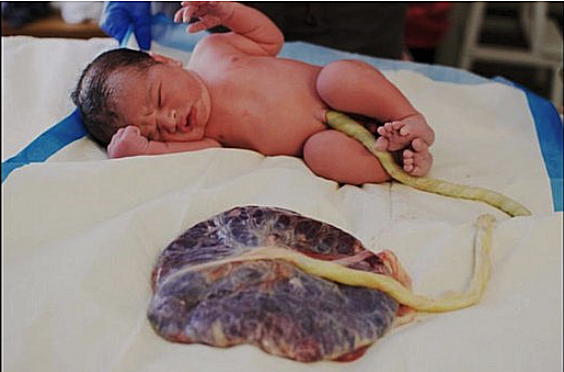 |
Finally, the the placenta and umbilical cord have delivered from the womb (Figure 31). When the umbilical cord is not cut, it naturally seals off about an hour after birth. The umbilical cord and attached placenta will fully detach from the baby anywhere from 2 to 10 days after birth. Modern practice ties off the cord tightly with heavy string after the cord has stopped pulsating. A clean shoelace, or sterile tape can be used about 4 inches from the baby; tie it again 2 to 4 inches from the first string, then cut between the two ties. It leaves a short piece of it, called a stump, attached to baby's belly button. The stump will naturally dry up and fall off within a few weeks after birth. |
Figure 31 Disposal of Umbilical Cord |

 |
The May 13, 2024 study on "Human embryos embrace asymmetry to form the body" shows that most of the human body forms from only one of the 2 cells in conception (see Figures 33,a, and 32 at "starting"). The research discovers that one of the two cells would become a fetus that divided faster later. The other one divides more slowly tended to turn into the yolk sac regressed eventually. |
Figure 33 Very Early Embryo [view large image] |
Another research paper published a month earlier on "Colonies of single-celled creatures could explain how embryos evolved" discovers the single-celled organism, which could develop into multicellular structures with remarkable similarities to embryos. |
 |
The research briefing article "Measuring the forces that shape early human embryos" reports that adequate surface tension is crucial for further embryonic development after the cleavage stage. Figure 34,a shows the degree of compaction as the C-M surface tension increase while the C-C's almost |
Figure 34 Surface Tension in Cleavage |
remains unchanged. The series of picutres on the top shows the change of compaction, i.e., C-M surface tension (in hours) during the experiment using a tiny glass tube (see top row in Figure 34,a). |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}