 , temperature T, pressure P, chemical potential
, temperature T, pressure P, chemical potential  , ... In equilibrium thermodynamics, the intensive properties are homogeneous, isotopic, and unchange through out the system. Such requirements are identified as :
, ... In equilibrium thermodynamics, the intensive properties are homogeneous, isotopic, and unchange through out the system. Such requirements are identified as :- Thermal Equilibrium - the temperature T is uniform through out the system (Figure 01,a).
- Mechanical Equilibrium - the pressure P is balanced by the external forces (Figure 01,b)
- Chemical Equilibrium - the chemical potentials for the various species remain constant, i.e., absence of any chemical reaction (Figure 01,c).
 |
As all these variables are held fixed at equilibrium, for example, the internal energy U can be expressed as : U = TS - PV + N ----- (1).
|
Figure 01 |
Such formulation with everything unchanged is not very useful in describing the changing world. Equilibrium thermodynamics gets around this problem by considering only the initial and final equilibrium states without regard to the intermediate details. |
 U = T
U = T


 (II
(II
 (
(
 0, there would be entropy dissipation even when
0, there would be entropy dissipation even when  0, (SII - SI) can be negative (driven by free energy infusion) as long as the inequality is satisfied. It means that orderly product can be created out of irreversible process at the expense of generating dissipative entropy
0, (SII - SI) can be negative (driven by free energy infusion) as long as the inequality is satisfied. It means that orderly product can be created out of irreversible process at the expense of generating dissipative entropy 



 n/
n/ ----- (7),
----- (7), 0, the entropy density s is a constant.
0, the entropy density s is a constant. , the whole system is at equilibrium
, the whole system is at equilibrium  .
. 0 ----- (9),
0 ----- (9),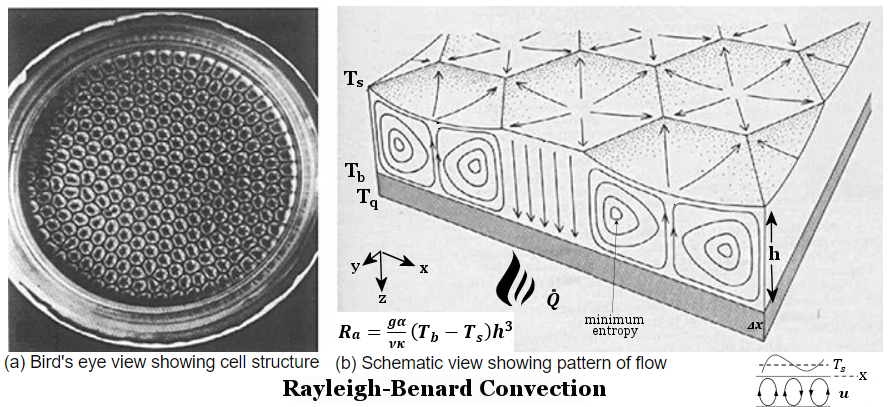



 dQ/T ----- (14),
dQ/T ----- (14),






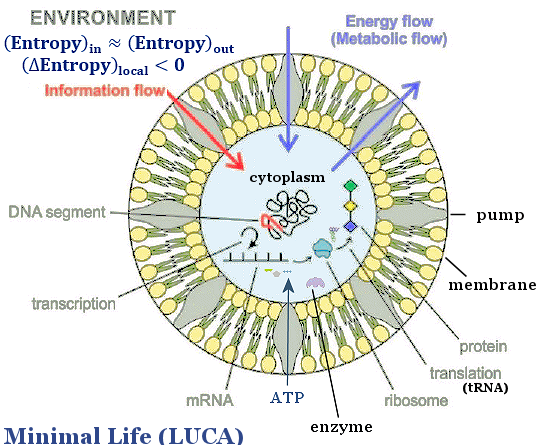
 m Energy Flow (erg/s-g)
m Energy Flow (erg/s-g)





 Oxygenic atmosphere
Oxygenic atmosphere
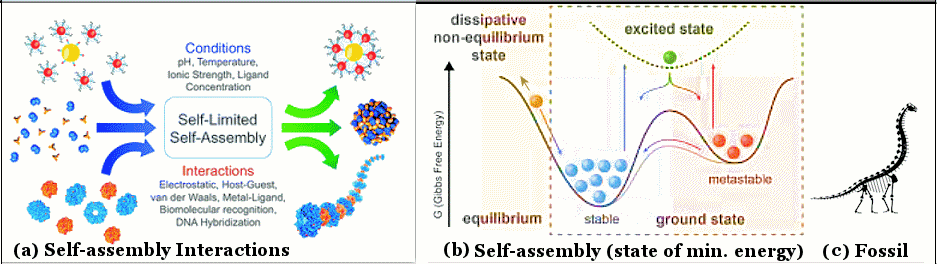
 can be calculated according to the formula shown in the Figure. This example illustrates different significance conveyed by entropy and information because they are related to
can be calculated according to the formula shown in the Figure. This example illustrates different significance conveyed by entropy and information because they are related to 


 (ni - 1), where N = 6.
(ni - 1), where N = 6.




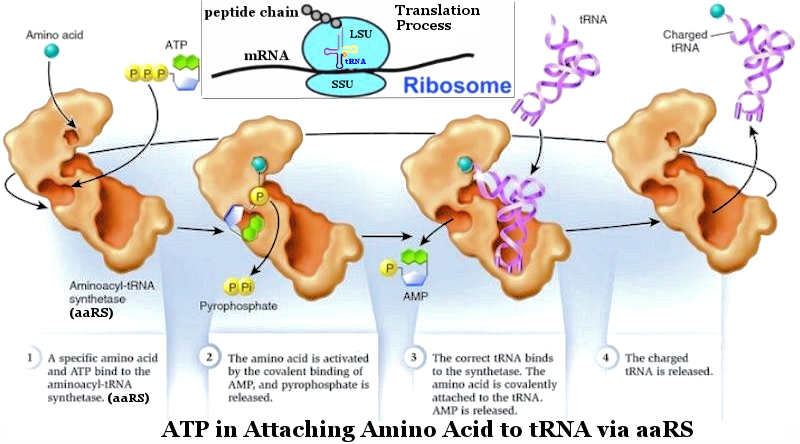

 ADP + Pi + 0.32 ev ----- (19).
ADP + Pi + 0.32 ev ----- (19).


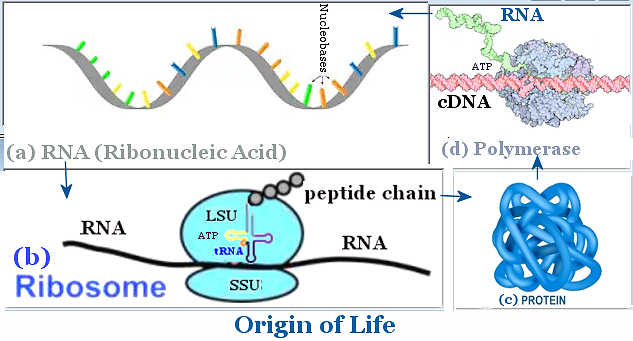
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![[view large image]](I15-39-)oLife.png){kind=link}